
Introduction
I. Background
I.a. From Geocentrism to the Expanding Universe
Briefly mention: Geocentrism → heliocentrism → 1 galaxy model → current known universe → multiverse theories
I.b. From Eukaryocentrism to the Tree of Life
Summarize: “Great chain of being” → 5 Kingdom system → 3 Domain system → Arcaryan hypothesis → Eocyte tree model → TACK hypothesis → Asgardian heritage
Then talk about: Endosymbiotic model of Eukaryogenesis
Also: The merging of Pongidae into Hominidae, introduction of Hominini, Gorillini, Hominina, and Panina, and the possible merging of Gorilla, Pan, and Homo.
I.b-1.) From Ancient Views to the 5 Kingdoms
| Animalia |
| Plantae |
| Fungi |
| Protista |
| Monera |
I.b-2.) From 5 Kingdoms to the Tripartite Tree
rRNA analysis showing closer evolutionary relationship between Eukaryotes and Archaea than between Archaea and Bacteria.
(Woese et al., 1990)
(Koonin, 2014)
| Eucarya |
| Archaea |
| Bacteria |
I.b-2α.) Annealing Model of the 3 Domains
Darwinian threshold, "crystallization" of the three primary lineages of life on Earth from out of the LGT-charactarized "pre-Darwinian" RNA world.
(Woese et al., 1990)
I.b-2α.) Arcaryan Hypothesis

I.b-3.) From 3 Domains to Symbiogenesis
For more information on the origin of mitochondria, please see my other article, "Mitochondrigeny": The Origin & The Evolution of The Mitochondrion, and Possible Implications for Astrobiology.
I.b-5.) From the Arcaryan Hypothesis to the Eocyte Tree
Eukaryotes within the Archaea, rather than the two as sister-groups.
I.b-5α.) TACK hypothesis
TACK (Thaumarchaeota-Aigarchaeota-Crenarchaeota-Korarchaeota) superphylum (Guy & Ettema, 2011)
I.b-5β.) Asgardian heritage
The hypothesis that the Eukarya evolved from within the so-called "Asgard" Archaeal superphylum.

Whereas 50 years ago, the Eukaryotes accounted for 4 of the 5 highest divisions of life on Earth, and 3 of those four were groups noted for containing large numbers of complex multicellular organisms, it now appears we Eukaryotes in general and complex obligately multicellular organisms in particular are nobbut a flimsy skin congealed atop the primordial soup, formed from the off-sloughings of the Tree of Life; an utterly insignificant twig on the expansive and diverse family tree that is Life on Earth.
I.c.) From Orthogenesis to Opportunistic Evolution
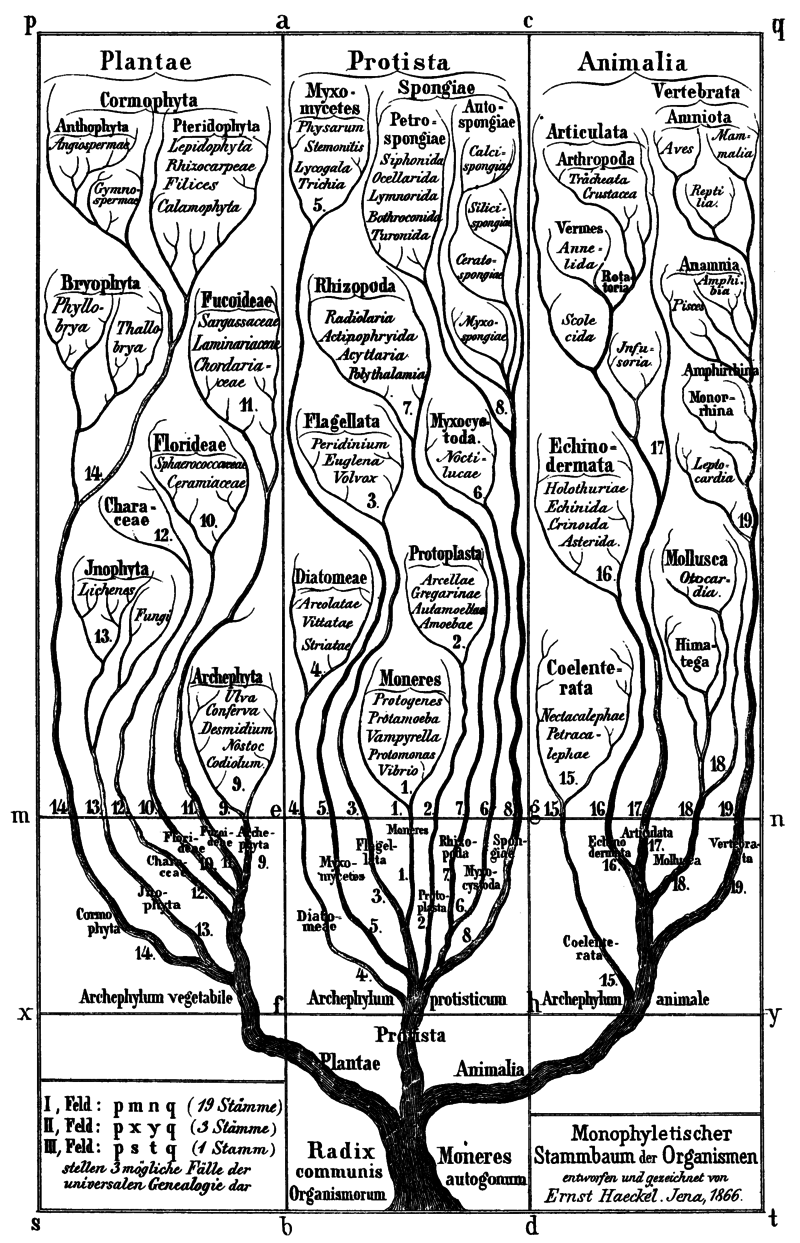
(Haeckel, 1866 C.E.)
Evolutionary scientists from the last decade of the 19th until around the mid-20th century commonly believed that evolution leads unidirestionally toward increasing complexity, in spite of a complete lack of empirical evidence in support of this view (McShea, 1991).
In the latter half of the 20th century, however, this began to change.
I.c-1.) Orthogenesis
Each generation of creatures is forged by those that were able to breed in the last, and in any generation individuals vary in fitness (Lucretius, 50 B.C.E.). This is plainly obvious for anyone to see; the artificial selection that humans have been practicing for millennia depends on it (Lucretius, 50 B.C.E.), this selfsame process occurred without the aid of humans during the “Tellurian age” (Lucretius, 50 B.C.E.), transforming the Earth over and over again (Lucretius, 50 B.C.E.), and the appearance of humans are a big part of the reason why the current wave (of artificially-selected domestic creatures and the few remaining wild species) replaced the last (Lucretius, 50 B.C.E.). Yet, strangely, people at various points in history have sought to mystify evolution, adding vaguely spiritual layers of meaning, often to bolster the human ego in a quasi-religious manner that places humans as the apex of some grand evolutionary process that drives things to become more complex over time (the implication here being that humans, a relatively "new" species, are the most complex form of life).
The belief that evolution leads purposefully toward greater complexity was held by John-Baptiste Lamarck, who believed that there was both a complexifying force and an adaptive force driving evolution. Most historical treatments of Lamarckianism focus on how Darwinism overturned Lamarck’s “acquired characteristic” model of the adaptive force, but neglect his complexifying force.
The term “orthogenesis” was first coined by Wilhelm Haacke (1893), and popularized by Theodor Eimer in (1898).
For a time this postulated internal drive toward evermore complicated organisms was suggested as an alternative to natural selection as the driving force behind evolution.
Proposed “internalist” (orthogenic) mechanisms for increasing complexity, according to McShea (1991), include:
1. Invisible fluids — Lamarck (1809) believed in the spontaneous generation of living things and that lineages tend to increase in complexity over time due to invisible fluids that lay ever-present in the environment but somehow become trapped inside organisms, modifying the cellular tissues through which they move, opening passages, forming canals, and finally creating new and different organs, enhancing complexity over time. Lamarck believed the effect of the environment, on the other hand, was mainly that of retarding this process of “complexification”. (Future theories of evolution would resurrect this idea of a “complexifying” force at odds with a simplifying environmental force, as it would later become apparent that natural selection, which eventually replaced Lamarck's “adaptice force”, tends to simplify and streamline organisms over time, while the “complexifying force” seems to be most prevalant during times of low selective pressure, suggesting that increasing complexity may be a thermodynamic process related to the accumulation of entropy when selective pressure is low enough to permit it; more on this later).
2. Instability of the homogeneous — Spencer (1890, in McShea, 1991) claimed that dynamic systems become less homogeneous and more concentrated as they evolve, referring to this as “The Law of Evolution” (note that here he was not referring to biological evolution, but to directional change in dynamic systems in general). In this conception, any perturbation to a homogeneous system leads to instability and re-organization of parts, resulting in complexity. Since life on Earth is in a constantly perturbed state (always metabolizing, respiring, &c.), it tends to become more complex over time.
3. Repetition and differentiation of parts — Cope (1871, in McShea, 1991) suggests that evolution occurs by acceleration of ontogenies (an idea first put forth by Haeckel) resulting in the repetition of existing parts which can then be differentiated, producing complexity.
4. The path of least resistance — Saunders & Ho (1976, 1981, in McShea 1991) posit that component additions are easier to achieve in development than deletions are because once evolved, components tend to be integrated into developmental pathways, making them difficult to remove (which we now know to be entirely wrong, see Maughan &al, 2007). According to Saunders & Ho, “[t]his asymmetry would be slight but sufficient to drive an evolutionary trend in complexity,” (McShea, 1991).
5. Complexity from entropy — In one version of this hypothesis, dynamic systems far from thermodynamic equilibrium may spontaneously develop complex structures and will tend to become more complex as those systems grow and age (Salthe 1985; Wicken 1987; in McShea, 1991). In another version, accumulation of entropy leads to morphological complexity, leading a configurational disorder that would typically be very harmful to a living organism, and increasing biological complexity is mainly a matter of increasing the fatal threshold (presumably via natural selection) for this configurational disorder.
I.c-2.) “Blind” Orthogenesis
It would be flattering to the scientific community to say that because natural selection could be observed to be the driving force behind evolutionary change in at least some cases (I’m sure we’re all familiar with the canonical textbook examples of industrial melanism, DDT-resistant insects, and antiobiotic-resistant bacteria), and could easily be inferred or deduced in others, orthogenesis obsolesced resultantly in accord with Occam’s razor. Unfortunately, this is not the case. The reality is that Ernst Mayr politically assassinated the term in the journal Nature in 1948 by stating that it implied "some supernatural force" (Ruse, 1996).
Whatever the reason, we defaulted back to the almost identical idea that life becomes more complex over time, not necessarily because of some mystical force that literally impels living things to become more complex over time, but because of a quasi-mystical principle that more complex things are inherently more fit and thus favored by natural selection. This was essentially an atheistic form of orthogenesis, and was the version of evolutionary theory championed by Darwin and Huxley.
While this “blind” or “naturalistic” form of orthogenesis may seem little different from the other, "purposed" variety of orthogenesis (that is to say, orthogenesis proper) from a modern perspective, one must understand that 20th century scientists were suffering from a mass delusion that complex multicellular lifeforms made up the bulk of life on Earth, regarded bacteria as little more than "germs", were unaware of the Archaea and artificially smashed them together with the bacteria into a kingdom called the “monera” while giving one Archaeal phylum, the Eukarya, four other kingdoms, then the highest taxonomic division of life, regarded the bacteria as “simple” and “primitive” without bothering to qualify or clarify the sort of complexity being spoken of or in regard to which qualities being compared, assumed without evidence that the earliest organisms should have been the simplest, that of course the modern organisms most distantly related to us bear the greatest resemblance to these primitive ancestors, did not typically regard the fossil record as containing anything of great interest until the Cambrian explosion about half a billion years ago, believed that there was a “great chain of being”-like evolutionary “March of Progress” toward ever-greater complexity, culminating in Earth's present biodiversity and Her youngest of species, such as our own, perched therefore at the end as the penultimate of complexity, a truth which expresses itself in that feature which at a cursory glance at Earth's biodiversity would at first seem to make humans most unique among animals: our grotesquely over-inflated brain. So married were they to this essentially orthogenetic narrative, that most scientists never thought to question it, arguing instead over why it seemed complexity had been increasing over time.
Proposed “externalist” mechanisms for increasing complexity, according to McShea (1991), include:
1. Selection for complexity — Rensch (1960) argues that the addition of parts permits more division of parts (occasional Drosophila mutants borne with legs growing where their antennae should be seem might be seen as aluding to something like this having occurred during the evolution of insects, however it must be remembered that modern insects and arachnids have fewer limbs and body segments than the more basal arthropods of yester-era), which makes more complex organisms more efficient, and this superior efficiency grants more complex creatures a selective advantage over simpler, less efficient organisms. Bonner (1988) expands on this by claiming that natural selection not only favors efficiency and therefore complexity, but also large size, which in turn may demand greater efficiency, which would also demand greater complexity.
2. Selection for other features — it may be that complexity increases passively as a side-effect of selection for other features, for example large size, which would generally necessitate a larger number of cells and hence create more opportunity for further differentiation of cell types.
3. Niche partititioning — Waddington (1969) suggests a system whereby increasing organismal diversity results in more complex ecological niches, which require greater organismal complexity to fill, which in turn creates more organismal diversity, and more complex niches, &c., in a sort of positive complexity feedback loop.
It must've seemed from their point of view that this “blind” or “naturalistic” orthogenesis could not have been more conceptually an opposite theory of evolution than orthogenesis proper. The version of the narrative invoking natural selection as an explanation during this time differed from orthogenetic theory merely in using as its mechenism a quasi-mystical principle that more complex organisms are inherently more fit and are therefore to be favored by natural selection, but to the average 20th century scientist, these two nigh-identic narratives were as different as night and day. Consider as an analogy how when the heliocentric model of the Solar system was first proposed, it must've seemed the polar opposite of geocentrism, yet compared to our modern point-of-view, a conception of the Universe placing a single star, our Sun, at the center of the known Universe, would seem little different from geocentrism; in both the case of heliocentrism and "blind" orthogenesis, scientists at the time were incapable of understanding how not-the-opposite of geocentrism and orthogenesis respectively their ideas really were, because the science simply hadn't evolved that far yet.
All of this is not to say that there were not during this time period skeptics of the idea that complexity had been gradually increasing over evolutionary time.
Though natural selection persists today as the most likely explanation for the majority of evolutionary change, the "blind" orthogenesis model of phylogeny that natural selection was intended as a mechanism for has since been discarded. Natural selection doesn't typically favor complexity, although it does arise occasionally (Ayala, 2007).
I.c-3.) Irreducible Complexity
Natural selection doesn't typically favor complexity, although occasionally complexity manages to arise nonetheless (Ayala, 2007).
While the term “irreducible complexity” generally refers to the creationist / theistic evolutionist hypothesis that certain biological systems could not logically have developed incrementally by means of natural selection as such would have required orthogenesis-like guided co-evolution of diverse, formerly independent components, an idea championed by the notorious Michael Behe (1996), there existed in the recent past an eerily similar notion in the scientific community: The idea that there is a certain “minimum” or “irreducible” complexity for life on Earth, that life started at the simplest, most irreducible level, and thus has become on average more complex over time not because evolution necessarily favors complexity, but simply because there is a limit to how simple things can become which means that any increase in the range of complexity of life on Earth would also necessarily raise the average complexity level. This concept was sound according to its own internal logic, but was based on 2 premises that were merely assumed rather than evidenced: [1] The idea that life on Earth has, on average, become more complex over time (McShea, 1991); and [2] the idea that once life had formed, nothing could evolve to become simpler than this “baseline” or “starting point” complexity.
Proposed “undirected” mechanisms for increasing complexity, according to McShea (1991), include:
1. Random walk — One possibility proposed by Fischer (1986) is that most evolutionary lineages could, due to sheer luck, just happen to wander in the direction of increasing complexity for no real reason at all.
2. Diffusion — Building upon this, Fischer proposes another possibility: If each evolutionary lineage follows its own random walk, and lineages tend to decrease in complexity as often as decrease, but there is a “complexity floor”, then the mean complexity for all lineages should climb upward over time. This is similar to Maynard Smith’s (1970) suggestion that if the first organisms had to be simple, then later organisms had “nowhere to go but up”.
3. The ratchet — Stebbins (1969) proposes that adaptive radiations resulting from the invasion of new habitats could cause major evolutionary jumps in complexity, not due to an intrisic drive toward complexity, but is contingent on pre-existing conditions such as promising morphological specializations. This lays the foundation for future jumps in complexity, resulting in the upper limit for complexity for life on Earth constantly ratcheting slowly upward.
I.c-4.) Back to Basics
Evolution is defined merely as a change in allele frequencies in a population over time. Evolution is most resoundingly not defined as an increase in complexity over time, and there's a damned good reason for that: If it were, it would’ve had to’ve been tossed out by now for lack of evidence.
Although as recently as 15-20 years ago, one could see statements that complexity has “obviously” increased over time (e.g., Carroll, 2001), this was already the viewpoint of a sad minority, obstinently digging their heels into the sand and proclaiming defiantly their refusal to go whithersoever the evidence lead, clinging instead to their eukaryocentric delusions of evolution leading inevitably to evermore complex forms of multicellularity, and their anthropocentric desire to situate humans, a relatively new species, at the end of a very long evolutionary march of progress. This was religion, not science. A position of faith utterly devoid of empirical support.
The idea of evolution driving toward ever-greater complexity is absurd on the face of it, when one considers that those who held this view also regarded the organisms who reproduce the fastest and have therefore experienced the greatest number of generations and hence have evolved the most since our common ancestor as the simplest and most primitive, whereas the organisms they regarded as the most complex (such has humans) tended to be reproductively lethargic and have therefore evolved far less since our common ancestor. That it took so long for the biological community at large to realize this should cause us all to hang our heads in shame for belonging to a species capable of such backward, delusional thinking.
“Since Nature banned with horror their increase,
And powerless were they to reach unto
The coveted flower of fair maturity,
Or to find aliment, or to intertwine
In works of Venus. For we see there must
Concur in life conditions manifold,
If life is ever by begetting life
To forge the generations one by one:
First, foods must be; and, next, a path whereby
The seeds of impregnation in the frame
May ooze, released from the members all;
Last, the possession of those instruments
Whereby the male with female can unite,
The one with other in mutual ravishments.”
— Lucretius, 50 B.C.E.
Natural selection cares nought for increasing complexity, but for survival and reproduction, as Lucretius evermore eloquently proclaimed in his poem, On the Nature of Things, more than two millennia ago. Often times, if not most of the time, it seems natural selection tends to simplify and streamline rather than to “complexify”. It tends to be the simpler systems, not the more complex ones, which are more efficient, as our bacterial brethren prove with their rapid reproductive rates and enviable adaptability (although part of what allows them to reproduce so fast, in addition to their simple morphology, is a doubly complex (compared to eukaryotes) system of transcription and translation at the heart of an incredibly advanced cellular metabolism). Complexity tends to arise when selective pressure is low, and may even represent an accumulation of entropy that would more likely’ve been trimmed away in a scenario with higher selective pressure in which efficiency would be more vital. This may help to explain why we see explosions in morphological/anatomical complexity immediately after mass extinction events, when we would expect selective pressure to be lowest.
While we can say with a high degree of certainty that there now exist on the Earth organisms far more morphologically complex than any that lived a billion years ago, and while it seems probable, though it is far from certain in light of recent theories regarding primal eukaryogenesis, that there were on Earth a billion years ago creatures more complex than anything that existed two billion years thither, which were, in turn, presumably, at least according to traditional and now it seems very possibly incorrect views regarding eukaryogenesis and abiogenesis, more complex than the first living things, it is not clear that this has been the overall evolutionary trend or that the cases of increasing complexity we've seen aren't actually the exception rather than the rule for evolution. Our perception that life has, on the whole, become more complex over time is now seen as possibly being the result of a sampling bias. It is now commonly believed that there has overall been no significant net-increase in the complexity of life on Earth, commonly understood that there is no particular reason to think that changes in allele frequencies over time would necessarily result in a gradual, unidirectional increase in complexity over time, commonly known that there are many examples of natural selection preferring to streamline and simplify than to keep unnecessarily complicated or to complicate further (especially being that complexity is evolutionarily costly), and commonly reasoned that in any case (whether there has been an overall net-increase, and overall net-decrease, or no significant overall change in the complexity of life on Earth), evolution (which is defined as a change in allele frequencies in a population over time) remains the only viable explanation for Earth's current biodiversity.
Much of this change in evolutionary thought has already filtered down into the popular culture. In the 2001 sci-fi comedy Evolution, for example, an ill-conceived attempt to use napalm to destroy an alien threat that reproduces and therefore evolves much faster in the presence of heat energy produces a gigantic amoeba-like organism, causing one character to remark, "That's evolution?!" to which another replies, "The 'simplest' organisms are often the most fit," which was a refreshingly accurate scientific observation in a major Hollywood production. Similarly, the articles Making Life Simple (Morton, 1999) and Evolution myths: Natural selection leads to ever greater complexity (Le Page, 2008) at New Scientist, as well as the Wikipedia article on the Evolution of biological complexity (Wikipedia, 2018) all reflect that evolution is no-longer seen as a unidirectional process that leads intrinsically to evermore morphologically complicated forms. That evolution is opportunistic and has no inherent preference for complexity is now so well-understood that to call it "academic" would be an understatement; that natural selection often favors simplicity and that organisms can vary not only in degree of complexity but in type (i.e., genomic complexity, morphological complexity, metabolic complexity, &c.), is now largely regarded as simply a matter of common sense.
I.d. From Anthropocentrism to the Hominid Family Tree
The merging of Pongidae into Hominidae
Introduction of Hominini, Gorillini, Hominina, and Panina
possible merging of Gorilla, Pan, and Homo.
I.e. From Human Exceptionalism to Modern Ethology
I.e-1.) Reasoning
I.e-2.) Social Complexity
I.e-3.) Language
Crows seem to have a complex language (Lorenz, Frings & Frings, Bannerman, and Chamberlain & Cornwell, in Powell, 1977)
I.e-4.) Tool Use
New Caledonian Crow, chimpanzees, gorillas, and orang-utans.
The New Caledonian Crow has been observed using and in some cases manufacturing its own tools.
I.e-5.) Construction
I.e-5α.) Bedding
Birds and great apes build nests.
I.e-5β.) Shelters
Moray eels, bears, moles, gophers, prairie dogs, beavers, wasps...
I.e-5γ.) Cities
Ants. Bees. Termites. Beavers.
I.e-6.) Space Travel
Microbe-containing impact ejecta from Earth may shield the colonies of microbial life within from deadly UV radiation while being small enough to be carried out to the Kuiper Belt via Solar winds, eventually to be deposited in the protoplanetary discs of future planetary systems as the Solar system moves through interstellar dust clouds in its course through the Milky Way (Wallis, 2003; Napier, 2003), seeding the Milky Way in just a few billion years, which, considering any Earth-like planet could hypothetically have done the same, makes it highly unlikely that Earth Herself was not a benefactor of such a process (Napier, 2003). Kind of makes humanity's quaint little trips to the Moon and back seem like... well, nothing even noteworthy. Though stepping foot on the Moon may have been a "giant leap" for mankind, it was infinitesimal compared to the untold hundreds of thousands of lightyears our bacterial brethren have likely spread themselves over by now.
I.e-7.) Specism
Not only do humans tend to regard their own breeding population as innately superior to all other organisms, and when asked to cite evidence, will generally simply proclaim that, "it's obvious". Some humans may think a bit more in-depth about the matter and recall things like language, the ability to reason, or the ability to make tools, and at first offer them as evidence of human superiority, but if it is pointed out that other creatures are capable of such thing, the human chauvinist will generally lapse back into, "what humans do is obvious different/superior; you can just tell!" That humans hold to the belief in the self-evident nature of their own superiority so ardently when there's so little empirical support for it, might be somewhat of a mystery, were it not for the explanatory power of the theory of evolution:
Very likely is it that most sexually reproducing organisms evolved instinctive behaviors and attitudes which prevent them from expending valuable resources on attempting to mate with organisms too genetically different from themselves for fruitful procreation to take place, or on attempting to compete with or empathizing with individuals too genetically different from themselves to have to worry about contending with for mates. Since most sexually reproducing organisms seem to select potential mates using some sort of superiority–inferiority scale (though the exact metric used to quantify superiority and inferiority may differ dramatically from one individual to the next, as evolution requires variation for natural selection to act upon), then it is very likely that for sexually reproducing organisms, sexual attraction and judgements of superiority (or at least adequacy) and inferior (inadequcy) are intimately linked, such that individuals one finds inferior or inadequate are not seen as potential mates, and vice versa: Individuals seen as unworthy of mating with (or competing with for mates) due to being too genetically different are probably perceived as qualitatively different forms of life and therefore reacted to as if inherently inferior. Undeniably this is what we observe among humans, and it seems nigh impossible to conclude that similar attitudes and behaviors are not the primary enabling factor behind virtually all "routine" interspecial conflict (predation, for example, or the scientific testing preformed by humans on non-human animals), or that an "overly conservative" version of these instinctive behaviors and attitudes isn't at the root of many examples of intraspecial conflict among humans (racism most especially, and possibly, albeit in a somewhat more derived form, also religious conflicts, homophobia, and classism more generally if we consider social behaviors, religious views, and social status as phenotypic traits that some humans might attempt to select for or against, in spite of any direct connection between these qualities and heritable genetic traits being tenuous at best).
It is vital to ever therefore remind ourselves that the impulse to regard our own breeding population as "special", "unique", "exceptional", "superior", or anything the like, is not objective reality but merely an animal extinct, intended to discourage unproductive sexual inclinations, expressed as a value judgement in terms of or akin to sexual selection (i.e., "inferior" versus "superior", "my type/kind" versus "not my type/kind"), and reinforced by selection bias and general self-delusion. Following naturally from this "humans outward", or, more to the evolutionary point, "our breeding population, outward" point of view, we instinctively in times past ranked the natural world in terms of usefulness to humans. Words for many common organisms are derived from anthropocentric utilitarian descriptions because before the advent of (ideally) unbiased scientific classification, humans were still fumbling about the natural world trying to classify things according to a selfish, infantile, "what does it do for me?" mindset, as evidenced in our own language by the Old English word deor (Modern English: "deer") originally referring to any wild game (ultimately from an Old High Germanic root, tior, meaning "beast", likely cognate with the Ancient Greek root *ther– for "beast" or "beastial", in words like "therian") while the word "cattle" comes from the Middle English catel, ultimately via Old North French from the Medieval Latin capitale for "wealth". The Christian Bible, which undoubtedly had a huge influence on European culture, says, "All flesh is not the same flesh: but there is one kind of flesh of men, another flesh of beasts, another of fishes, and another of birds," (I Corinthians 15:39 KJV) which, when one considers how the Christian Bible uses the word "flesh", and that bats are classified as birds and whales as fish in the book of Leviticus, are clearly disgnations of function, behavior, or inclination rather than biological categories. Plants were not even considered alive under this system, and humans, a single species, represented one quarter of the four highest-level classifications of living things. Later we alotted four of the five highest divisions of life on Earth to members of our own obscure little phylum of Archaea while artificially and eukaryocentrically smashing all of our fellow Archaea together with the Bacteria into a single kingdom! We once believed that evolution predicts ever-increasing morphological complexity simply because that was the trend we thought we saw when in the 20th century we looked to the fossil record while anthropocentrically placing greatest emphasis on those organisms which we found most familiar (vertebrates over invertebrates, animals over plants and fungi, and multicellular forms over unicellular or sub-cellular forms, &c.), a perception based ultimately not in evidence but in an obsessive desire to believe our own relatively young species to be the latest in a march of progress toward ever-increasing complexity. We once believed that our own breeding population was the only that could reason, or that possessed language, as an extension of the religious belief that only creatures of our breeding population have souls. We held to these default assumptions with such vigor, and resisted so adamantly Nature's rebuke as at every step the process of science dragged us out into the light and exposed us with each iteration evermore for the insignificant cell colonies that we are, not because these things were evidenced by facts borne out of Nature Herself, nor due in any way whatever to objective truths of any kind, but because of an utterly delusional sense of specism; an obsession with our own breeding population (and thus ultimately a very biological obsession with sex) justified only from a utilitarian perspective by evolutionary necessity.
That we consider ourselves to be so special, is proof of just how un-special we are.
II. Part Two
II.a. From Geocentrism to the Expanding Universe
How we determined the Sun does not revolve around the Earth, and how we learned our galaxy was not the only galaxy, and why some physicists think there might be a multiverse (cosmic inflation & Everett’s interpretation of quantum physics)
II.a-1.) From Geocentrism to Heliocentrism
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.b. From Eukaryocentrism to the Tree of Life
How we transitioned from a “great chain of being” model to a branching tree of “increasing complexity” with “higher” lifeforms distinguished from “lower” or “simpler” lifeforms, with the so-called “monera” (Bacteria+Archaea but lacking Eukaryota, essentially) being regarded as primitive relics of a bygone era before we “advanced” eukaryotes sprang up from them, or some such nonsense, to the realization that the Eukarya are truly but a small and insignificant branch on a great and diverse tree of life.
II.b-1.) From the Ancient Views to the 5 Kingdoms
New Concepts of Kingdoms
of Organisms
R. H. Whittaker. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms.
http://www.ib.usp.br/inter/0410113/downloads/Whittaker_1969.pdf
II.b-2.) From 5 Kingdoms to 3 Domains
rRNA analysis showing closer evolutionary relationship between Eukaryotes and Archaea than between Archaea and Bacteria.
(Woese et al., 1990)
(Koonin, 2014)
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.b-2α.) Annealing Model of the 3 Domains
Darwinian threshold, "crystallization" of the three primary lineages of life on Earth from out of the LGT-charactarized "pre-Darwinian" RNA world.
(Woese et al., 1990)
II.b-2α.) Arcaryan Hypothesis
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.b-3.) From 3 Domains to Symbiogenesis
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
For more information on the origin of mitochondria, please see my other article, "Mitochondrigeny": The Origin & The Evolution of The Mitochondrion, and Possible Implications for Astrobiology.
II.b-5.) From the Arcaryan Hypothesis to the Eocyte Tree
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.b-5α.) TACK hypothesis
TACK (Thaumarchaeota-Aigarchaeota-Crenarchaeota-Korarchaeota) superphylum (Guy & Ettema, 2011)
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.b-5β.) Asgardian heritage
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.c.) From Orthogenesis to Opportunistic Evolution
II.c-1.) Orthogenesis
While orthogenesis might seem relatively easy to debunk from a modern point of view, it was not so when the only alternative being offered was a version of evolutionary theory that, like orthogenesis, posited that complexity should increase over time.
Could we have proven orthogenesis wrong with any late 19th century to mid-20th century knowledge? As it turns out, yes, we easily could have! In “The Origin of Species”, Darwin conjectures that when natural selection doesn’t work (i.e., when selective pressure is extremely low), organisms will remain unchanged for long periods of time (Seravin, 2001).
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.c-2.) “Blind&rdquo Orthogenesis
Since this conception of evolution for the most part made all of the same predictions as the alternative model of evolutionary theory was making at the time, orthogenesis could not be officially “debunked” (aside from the aforementioned “living fossils”) without also shedding doubt as to whether evolution truly proceeded as we had always envisioned. It therefore took multiple examples of reduction (evolutionary processes that tend to reduce complexity over time, such as phenotypic loss, neoteny, and parasitism) for biologists to realize that evolution is not necessarily about organisms becoming more complex over time.
Parasitic organisms such as Plasmodia and mycoplasmas are reductionist lifeforms that have dispensed with a number of traits that parasitism has made obsolete in their lineages (Sirand-Pugnet &al., 2007).
Phenotypic loss describes morphological simplification and streamlining over time (Maughan &al, 2007). Modern sharks have for the most part adopted a generic “shark shape” that while now familiar is a much simplified version of the morphologically diverse and often intricately morphologically complex group from which they evolved. Consider for example ancient sharks like the 420 MYO Mongolepis, 390-320 MYO Stethacanthus, the 365 MYO Cladoselache, the 360 MYO Falcatus, the 350 MYO Edestus, the 270 MYO Helicoprion, the 259-66 MYO Hybodus, the 200 MYO Xenacanthus, or the 120-70 MYO Scapanorhuynchus. Modern cephalopods also show evidence of simplification and phenotypic loss compared to ancient cephalopods, many of whom had incredibly elaborate shell designs. Similarly, slugs evolved from snails, not the other way around. Modern jellies have likewise been much simplified compared to the ancestral group out of which they evolved.
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
Although natural selection certainly works as an explanation of evolutionary change over time, there are enough instances of reduction (far too many to create a comprehensive list here) that we either needed to re-evaluate whether natural selection tends to favor complexity, or we needed to re-examine whether many of the evolutionary changes documented in the fossil record were due primarily to natural selection or possibly due primarily to some other force, such as a thermodynamic entropy-driven model for increasing complexity, or possibly some form of genetic drift in low-selective-pressure situations (“constructive neutral evolution”). Though this issue is far from settled as of the year 2018 C.E., it currently appears as though both of these alternatives to “blind” orthogenesis have found a great deal of support in the past few decades, and are far from mutually exclusive.
II.c-3.) Irreducible Complexity
Whether or not this “irreducible complexity” model of evolution is more correct than the previous orthogenesis and orthogenesis-like version of evolution via natural selection can be tested by comparing predictions with the distribution of complexity among the biota.


|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
It’s also possible that there‘s been a passive trend toward increasing complexity (Carroll, 2001).

|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
II.c-3α.) Consideration of Viruses, Organelles, and Other "Pseudobiota"
Viruses, organisms so simple that there is some debate over whether they should qualify as living things, are the dominant biological entities on this planet (Koonin & Wolf, 2012).
There are three main hypotheses for the viral origins:
1. Virus-first hypothesis — viruses could have evolved from complex molecules of protein and nucleic acid before cells first appeared on earth (Villarreal, 2005) and had a profound influence on the evolution of these first cells (Nasir &al., 2012). This idea has been criticized for contradicting the definition of the word “virus”, which implies requiring a host cell for replication (if viruses came first then there were no host cells). Nevertheless, it is possible that small, virus-like replicons which were not yet parasites are the direct ancestors of at least some modern viruses.
2. Reduction hypothesis — viruses could be reduced from prokaryotic organisms such as the Rickettsia (whence our mitochondria also derive) or Rickettsia-like endoparasites (Nasir &al., 2012; Koonin & Wolf, 2012).
3. Escape hypothesis — viruses may have begun as segments of their host genomes (Nasir &al., 2012; Koonin & Wolf, 2012), and one may suspect perhaps even from bacterial and archaeal plasmids, such as for example Rickettsial plasmids.
Contemporary virologists hypothesize that no single one of these hypotheses is “the” definitive explanation for the evolution of viruses, but perhaps that a combination of all three occurred, resulting in a polyphyletic grouping (Nasir &al., 2012; Koonin & Wolf, 2012; Forterre, 2014).
Since viruses lack the ability to reproduce on their own, it is fair at least to strongly suspect that they are simpler than the first self-replicating organisms would have been. This creates a problem for the notion that life began at or near the simplest, most irreducible level of complexity possible.
If we assume a baseline complexity level roughly comparable to modern prokaryotes (there are reasons for thinking the first living things may have been either more or less complex than modern prokaryotes; discussed later), then determining whether our perception of life on Earth as having generally evolved in the direction of higher complexity is an example of selection bias due to our eukaryocentrism, or whether the idea of a quasi-mystical "drive toward complexity" has merit, is simply a matter of comparing the number of times complex multicellularity has evolved, minus the number of reversions to unicellularity, compared to the number of times viruses, prokaryote-derived organelles like mitochondria and chloroplasts, and other "quasi-biota" or "pseudobiota" have evolved.
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
Numbers for this analysis were difficult to come by. However, consider the following:
N1: Multicellularity has evolved at least 46 times in eukaryotes (Grosberg & Strathmann, 2007): At least once (perhaps twice) in animals, three times in fungi, and repeatedly in Chloroplastids (algae and land plants).
N2: a cursory search through the literature did not immediately reveal any estimates regarding the total known or estimated number of reversions to unicellularity, however some examples include Saccharomyces and many other related yeasts in the Saccharomycotina (Medina &al, 2003), Cryptococcus albidus and related species (Medina &al, 2003), yeasts in the Taphrinomycotina (Medina &al, 2003), Schizosaccharomyces sp. (Medina &al, 2003), and Pneumocystis carinii (Medina &al, 2003). While yeasts have long been regarded as secondarily unicellular (Whittaker, 1969), they belong to the phyla Ascomycota (sac fungi) and Basidiomycota (mushrooms, puffballs, stinkhorns, bracket fungi, other polypores, chanterelles, boletes, jelly fungi, earth stars, smuts, rusts, bunts, mirror yeasts, and the aforementioned human pathogenic yeast Cryptococcus) within subkingdom Dikarya (“higher” fungi), which means the grouping is informal and polyphyletic; the yeasts taxonomically and cladistically nest within otherwise “complex” multicellular taxa and therefore most likely represent several independent reversions to unicellularity. That this is generally the preferred assumption to the idea of yeasts being more primitive and their various multicellular relatives as more derived has as much to do with genome size as it does with Occam’s razor: positing large-scale reduction in numerous independent lineages requires fewer unproved or unprovable assumptions than imagining large-scale convergent evolution toward multicellularity in a variety of lineages, many of which end up coincidentally or through some unseen lateral gene transfer end up sharing genes related to multicellularity. That reduction is the evolutionarily easier explanation here is very telling about whether we should expect in general for evolution to tend toward greater complexity. In contrast to these multiple reversions to unicellularity,
N3: Viruses are now generally assumed to be a polyphyletic group that has evolved from the eukarya and prokarya both through reduction and segregation (Nasir &al., 2012; Koonin & Wolf, 2012), as well as from ancestors that were co-evolving with the first cells. While we’ve yet to definitively work out exactly how many times viruses have evolved from cellular lifeforms, the number of independent evolutions here is likely immense.
N4: The only organelles known for certain to have evolved from prokaryotes are plastids and mitochondria, however a variety of other organelles, such as ribosomes and prokaryotic plasmids (which are not true organelles but share many features in common with them), may have also begun their evolutionary lineages as cellular organisms.
So while it appears as though reversions to unicellularity from multicellularity are relatively common, that a likely huge number of viruses have evolved from prokaryotic and eukaryotic organisms through both segregation and reduction, and that at least two and possibly a great deal more organelles evolved from prokaryotes, and that plasmids may have evolved from prokaryotes multiples different times, meaning that it is relatively obvious that what authors have in the past called “regressive” evolution is actually far more common than orthogenesis-like “progressive” evolution, lack of exact numbers here makes it impossible, at the present moment, to quantify exactly by what proportion “regressive” evolution seems to be preferred.
Another approach to this problem might be to measure the evolutionary success of these strategies not in terms of how many times “regressive” or “progressive” evolution has taken place with regard to unicellularity and multicellularity, but the relative success of the different strategies. For this we would still assume the same roughly prokaryote-equivalent “baseline” complexity (again, the first living things and/or Last Universal Common Ancestor may have been either more or less anatomically complex than modern prokaryotes, and there are good reasons for supporting either scenario), we would merely have to compare the number of multicellular organisms, unicellular organism, and “pseudobiota”.
Thus, failure to acknowledge the pseudobiota is in this context a fallacy of a limited sample. While some might object to the consideration of the pseudobiota with regard to the question of complexity on the grounds that viruses fail to fulfill all the criteria for being considered "alive" and thus are not examples of “life”, to maintain that there is a certain “minimum” or “irreducible” complexity that living things never fall below while regarding any biological entity which evolves below this invisible line of irreducible complexity as "not alive", is fallacious reasoning; it is a tautology designed to justify selection bias, or in other words: Refusal to acknowledge the pseudobiota is in this context a No-True-Scotsman argument.
II.c-3β.) Uncertain Baseline Complexity
Primal Eukaryogenesis: On the Communal Nature of Precellular States, Ancestral to Modern Life (Egel, 2011)
The Nuclear Compartment Commonality Hypothesis (Staley, 2013)
|
|
|||||
| ↓ | ↓ | |||||
|
|
|||||
| ↓ | ↓ | |||||
|
Compare ⇄ |
|
Note that if the Eukarya are as deeply nested within the Archaea as currently appears to be the case (Eme &al, 2017), then one of the consequences of the NuCom hypothesis being true is that a number of independent lineages leading to all extant non-eukaryotic Archaea must've shed their nuclear compartments independently of one another, so that while eukaryogenesis only occured once, and much earlier than traditionally hypothesized, "prokaryogenesis" has occured many times over, which is consistent with phenotypic loss being evolutionarily easier than the addition of new and complex traits.
This, however, only gets us as far back as the Last Universal Common Ancestor (LUCA), the common ancestor of the Eukarya/Archaea and the Bacteria. Even if the evolutionary trend has been toward greater simplicity rather than toward greater complexity since the LUCA, what of the First Universal Common Ancestor (FUCA), or first lifeforms? Wouldn’t a proto-eukaryotic LUCA still be far more complex than the very earliest lifeforms? Not necessarily! If we define the first living things as the first biological entities made up of nucleic acids housed in a lipid bilayer, then the Primal Eukaryogenesis hypothesis (Egel, 2011) suggests the first living things may have been more complex than
II.c-4.) Moving Forward
The relevant question now is why past generations of scientists wrongly inferred an overall, over-arching, largest-possible-scale evolutionary trend toward ever-increasing complexity since the origin of life, despite a complete lack of evidence.
Reasons suggested by McShea (1991) for all of this “gestalt” include:
1. Humans are essentially psychic — “One [possibility] is that complexity does increase, and that we unconsciously compute complexity between earlier and later organisms with some innate, cognitive algorithm, or even perceive differences directly, in ways that we simply cannot yet articulate. If so, then our only project is to discover how to say what we already know,” (McShea, 1991). This is probably the least likely of all scenarios, however it appears to be the assumption that the majority of scientists during the late 19th and early-to-mid 20th were operating under.
2. Properties other than complexity are causing the gestalt — as McShea writes, “Comparing a cat with a clam, for example, many will get a vague impression that ‘something more’ is going on in the cat.” McShea then goes onto question whether this “something more” is actually greater complexity, or other qualities like greater intelligence, greater mobility, or simply greater similarity to humans. McShea then reminds that complexity has to do with number of different kinds of parts and the irregularity of their arrangement. McShea points out that since cats and clams are so anatomically different from one another, comparing their parts is not straight-forward, and that if cats strike us as seeming to have more parts, it could simply be that they are, on average, larger creatures and much more closely related to us, making their parts easier for us to identify (both in being easier for us to see and in being more familiar to us). “Possibly, as one reviewer suggested, organisms simply look more and more different from modern ones as we scan further and further back in time; if the moderns are assumed to be very complex, then less familiar might be mistaken for less complex. The human perspective, like any other, has its biases.” (McShea, 1991)
3. Hyperbole of unrepresentative samples — it’s also possibly that the few clearcut cases of increasing complexity (such as the rise of multicellularity) so dominate our perception of evolution that we wrongly generalize such cases to long-term trends. This especially might be skewing our perception of how evolution works because such leaps forward in anatomical complexity might not be due to “day-to-day” evolutionary forces like as genetic drift or natural selection, but due instead to exceptional, out-of-context events like meteor strikes (for example, mitochondrial acquisition could never have happened without bacteria, who might not even be native to Earth (Scott, 2018)!). Those who held to the belief that complexity tends to increase over time were thus living in the exceptions.
4. Projection of technological trends — just as it is popularly imagined that the evolution of tool-manufacturing has instilled in our minds the false “common sense” notion that all useful things require a creator, the trend in technology toward increasing complexity of devices may be causing us to read a similar trend into the history of life on Earth. These “common sense” notions, or “obvious truths” may simply be the part of our brain that knows how to recognize a human-created artifact trying to find such patterns even where none exist (not wholey unlike a religious person thinking he or she has seen the face of the Virgin Mary in a piece of toast).
5. Conflation of complexity with progress —
II.e. From Human Exceptionalism to Modern Ethology
In Aesop’s famous fable, “The Crow and the Pitcher”, a thirsty crow drops pebbles into an under filled pitcher of water, raising the water level high enough for the bird to drink. Recent scientific research has corroborated this tale, showing that, faced with a similar situation, crows will drop stones into a tube containing water, but not into a tube containing sand, and will choose to drop solid objects into the water rather than hollow ones (which demonstrates an understanding of buoyant objects being of limited value for water displacement), revealing that crows have a basic understanding of cause and effect that human children struggle with until age 5 (Castro & Wasserman, 2015; still tracking down Wasserman's original paper).
II.e-1.) Reasoning
II.e-1α.) Theory of Mind
Crows and chimpanzees
II.e-1β.) Cause & Effect
In Aesop’s famous fable, “The Crow and the Pitcher”, a thirsty crow drops pebbles into an under filled pitcher of water, raising the water level high enough for the bird to drink. Recent scientific research has corroborated this tale, showing that, faced with a similar situation, crows will drop stones into a tube containing water, but not into a tube containing sand, and will choose to drop solid objects into the water rather than hollow ones (which demonstrates an understanding of buoyant objects being of limited value for water displacement), revealing that crows have a basic understanding of cause and effect that human children struggle with until age 5 (Castro & Wasserman, 2015; still tracking down Wasserman's original paper).
II.e-1γ.) Symbolism
Crows have been proven to be able to understand symbolism (Castro & Wasserman, 2015; still tracking down Wasserman's original paper).
II.e-1γ.) Science & Deduction
Crows perform their own version of forensic anthropology, gathering around corpses of their fallen comrades and attempting to determine cause of death as a sort of risk-assessment, and possibly also as a form of mourning or funerary rite, though the latter is difficult to validate scientifically (Swift, 2015).
II.e-1δ.) Analogy
Crows
II.e-2.) Social Complexity
II.e-3.) Language
II.e-4.) Tool Use
II.e-5.) Construction
II.e-6.) Space Travel
II.e-7.) Specism
References
Francisco J. Ayala. Darwin's greatest discovery: Design without designer. Proceedings of the National Academy of Sciences May 2007, 104 (suppl 1) 8567-8573; DOI: 10.1073/pnas.0701072104
http://www.pnas.org/content/104/suppl_1/8567.full
Sean B. Carroll. Chance and necessity: the evolution of morphological complexity and diversity. Nature 409, 1102–1109 (22 February 2001). doi:10.1038/35059227
https://www.nature.com/articles/35059227
Castro, Leyre & Wasserman, Ed. “Crows Understand Analogies” Scientific American (February 10, 2015)
<https://www.scientificamerican.com/article/crows-understand-analogies/>
Richard Egel. Primal Eukaryogenesis: On the Communal Nature of Precellular States, Ancestral to Modern Life. Life 2012, 2(1), 170-212; Received: 8 November 2011; in revised form: 29 December 2011 / Accepted: 11 January 2012 / Published: 23 January 2012 doi:10.3390/life2010170
http://www.mdpi.com/2075-1729/2/1/170/htm
Laura Eme, Anja Spang, Jonathan Lombard, Courtney W. Stairs & Thijs J. G. Ettema. Archaea and the origin of eukaryotes. Nature Reviews Microbiology volume 15, pages 711–723. (2017) doi:10.1038/nrmicro.2017.133 Published: 10 November 2017, Corrigendum: 27 November 2017, Corrected online 27 November 2017
https://www.nature.com/articles/nrmicro.2017.133
Guy L1, Ettema TJ. The archaeal 'TACK' superphylum and the origin of eukaryotes. Trends Microbiol. 2011 Dec;19(12):580-7 Epub 2011 Oct 20. doi: 10.1016/j.tim.2011.09.002
https://www.ncbi.nlm.nih.gov/pubmed/22018741/
Koonin, Eugene V. “Carl Woese's vision of cellular evolution and the domains of life.” RNA Biology. v.11(3); 2014 Mar 1 PMC 4008548, 2014 Mar 1; 11(3): 197–204. Published online 2014 Jan 16. doi: 10.4161/rna.27673
<https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4008548/>
Koonin, Eugene V., and Wolf, Yuri I. “Evolution of microbes and viruses: a paradigm shift in evolutionary biology?” Front Cell Infect Microbiol. 2012; 2: 119. Published online 2012 Sep 13. doi: 10.3389/fcimb.2012.00119
<https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3440604/>
Le Page, Michael. Evolution myths: Natural selection leads to ever greater complexity. New Scientist. Daily News 16 April 2008
<https://www.newscientist.com/article/dn13617-evolution-myths-natural-selection-leads-to-ever-greater-complexity/>
Lucretius, On the Nature of Things, 50 B.C.E., in Understanding Evolution: History, Theory, Evidence, and Implications by R. G. Price - March 5, 2006; Updated - May 2, 2006, http://www.rationalrevolution.net/articles/understanding_evolution.htm
Maughan, H., Masel, J., Birky, C. W., & Nicholson, W. L. (2007). The Roles of Mutation Accumulation and Selection in Loss of Sporulation in Experimental Populations of Bacillus subtilis. Genetics, 177(2), 937–948. http://doi.org/10.1534/genetics.107.075663
McShea, D.W. Complexity and evolution: What everybody knows. Biology & Philosophy. Volume 6, Issue 3, pp 303–324 (1991). https://doi.org/10.1007/BF00132234
<https://link.springer.com/article/10.1007%2FBF00132234#citeas>
Morton, Oliver. Making Life Simple. New Scientist. Feature 16 January 1999
<https://link.springer.com/article/10.1007%2FBF00132234#citeas>
Napier, W. M. A mechanism for interstellar panspermia. Mon. Not. R. Astron. Soc. 348, 52-61 (2004). Accepted 2003 October 2. Recieved 2003 October 1; in original form 2003 April 4. doi:10.1111/j.1365-2966.2004.07287.x
http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2966.2004.07287.x/abstract
Oren, A. (2004). "Prokaryote diversity and taxonomy: current status and future challenges". Philos. Trans. R. Soc. Lond. B Biol. Sci. 359 (1444): 623–38. doi:10.1098/rstb.2003.1458.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1693353/
Powell, Robert W. “Operant Conditioning in the Common Crow (Corvus brachyrhynchos).” The Auk (Oct., 1972), vol. 89, no. 4 pp. 738-742
<https://www.jstor.org/stable/4084106?seq=1#page_scan_tab_contents>
Schloss, P.; Handelsman, J. (2004). "Status of the microbial census". Microbiol Mol Biol Rev. 68 (4): 686–91.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC539005/
Scott, D. J. (2018). "Mitochondrigeny": The Origin & The Evolution of The Mitochondrion, and Possible Implications for Astrobiology. D. J. Scott’s Website. Our Family Tree.
https://djonscott.com/m-geny.html
Sirand-Pugnet, P., Lartigue, C., Marenda, M., Jacob, D., Barré, A., Barbe, V., … Citti, C. (2007). Being Pathogenic, Plastic, and Sexual while Living with a Nearly Minimal Bacterial Genome. PLoS Genetics, 3(5), e75. http://doi.org/10.1371/journal.pgen.0030075
James T Staley. The Nuclear Compartment Commonality Hypothesis, Enucleation and the Evolution of the Bacteria and Eukarya. Journal of Astrobiology & Outreach, 1:3 2013. DOI: 10.4172/2332-2519.1000105
https://www.omicsonline.org/open-access/the-nuclear-compartment-commonality-hypothesis-enucleation-and-the-evolution-of-the-bacteria-and-eukarya-2332-2519.1000105.pdf
Swift, Kaeli. “Why crows gather around their dead” Corvid Research (September 26, 2015)
<https://corvidresearch.blog/2015/09/26/why-crows-gather-around-their-dead/>
Wallis, Max K. and Wickramasinghe, N. C. Interstellar transfer of planetary microbiota. Mon. Not. R. Astron. Soc. 348, 52-61 (2004). Accepted 2003 October 23. Recieved 2003 October 23; in original form 2003 June 5. doi:10.1111/j.1365-292004.07355.x
https://www.researchgate.net/publication/227621140_Interstellar_transfer_of_planetary_microbiota
Whitman WB, Coleman DC, Wiebe WJ (June 1998). "Prokaryotes: the unseen majority". Proceedings of the National Academy of Sciences of the United States of America. 95 (12): 6578–83.
http://www.pnas.org/content/pnas/95/12/6578.full.pdf
Proc. Nati. Acad. Sci. USA Vol. 87, pp. 4576-4579, June 1990
Evolution Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eukarya
CARL R. WOESE, OTTO KANDLER, AND MARK L. WHEELIS. Contributed by Carl R. Woese, March 26, 1990
<http://www.pnas.org/content/87/12/4576>
C R Woese and G E Fox. Phylogenetic structure of the prokaryotic domain: the primary kingdoms.
<http://www.pnas.org/content/87/12/4576>